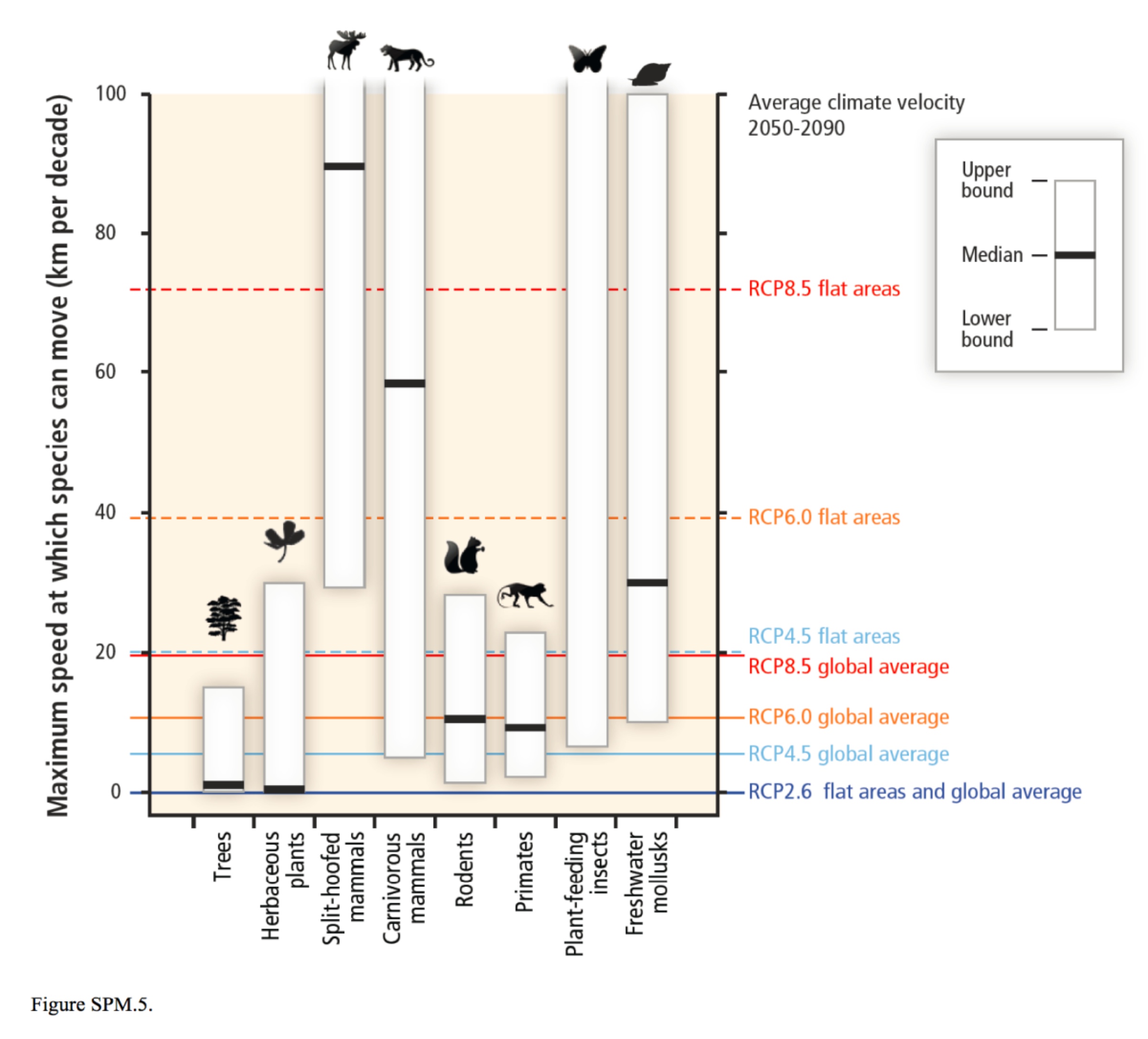
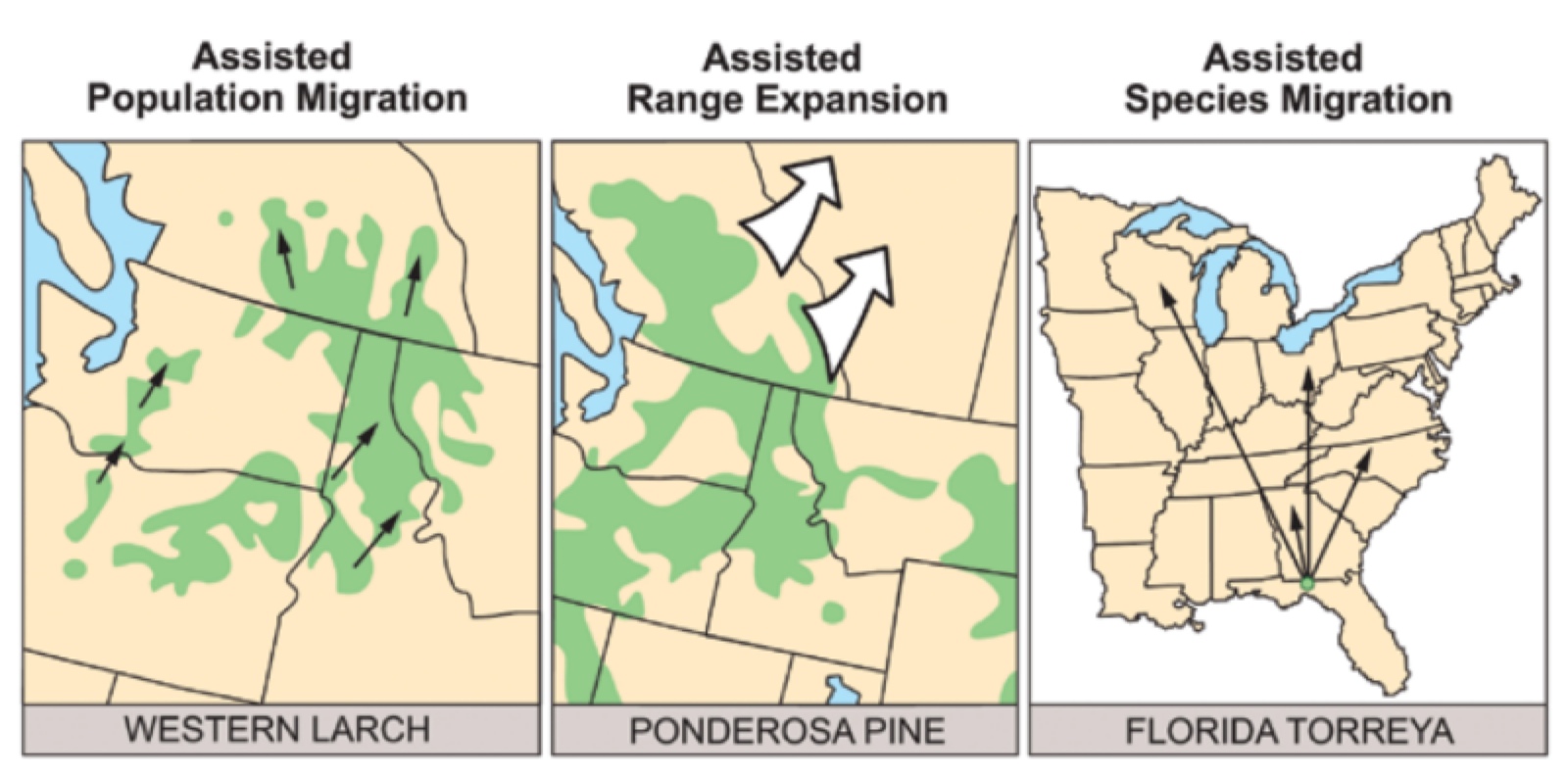
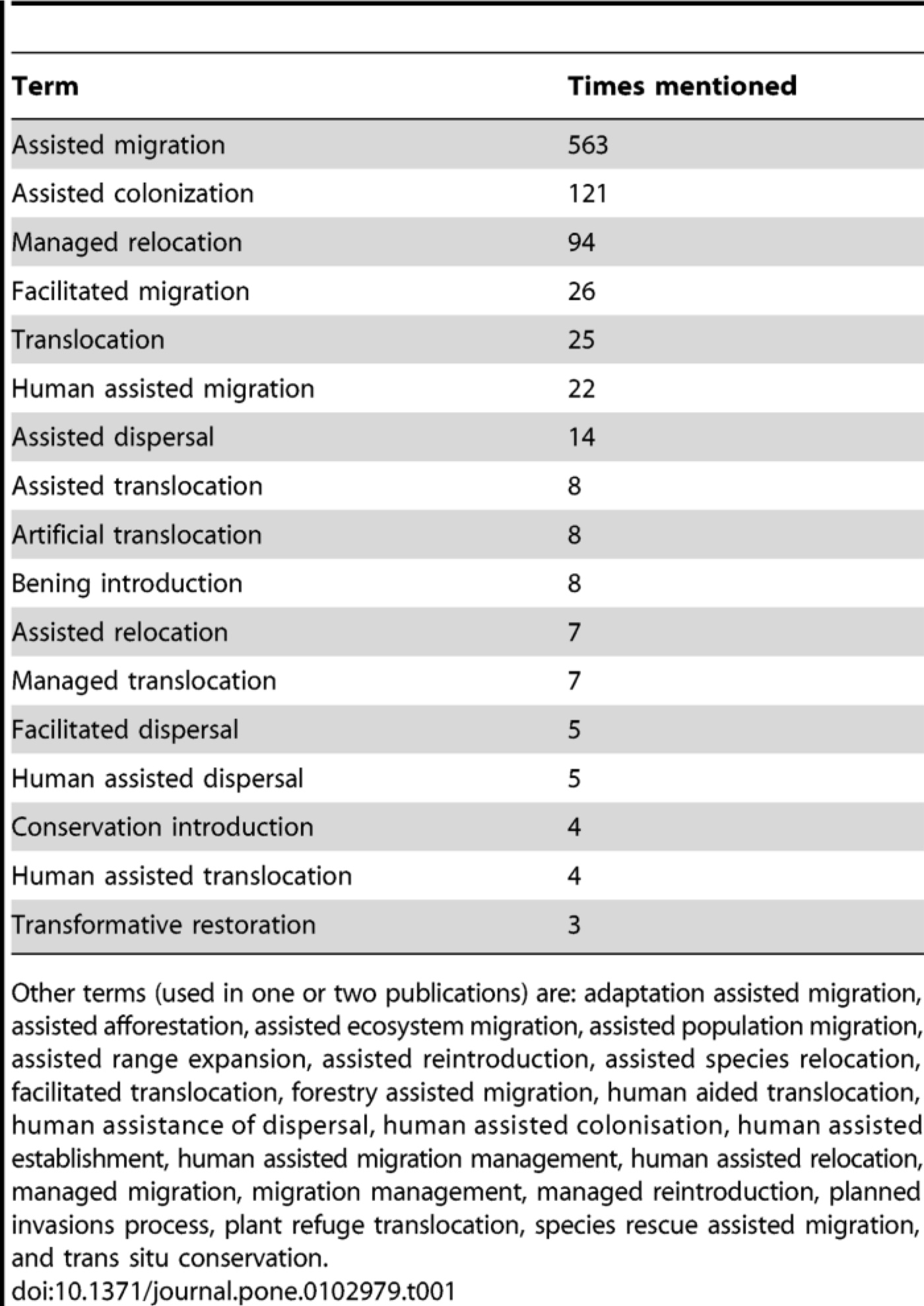
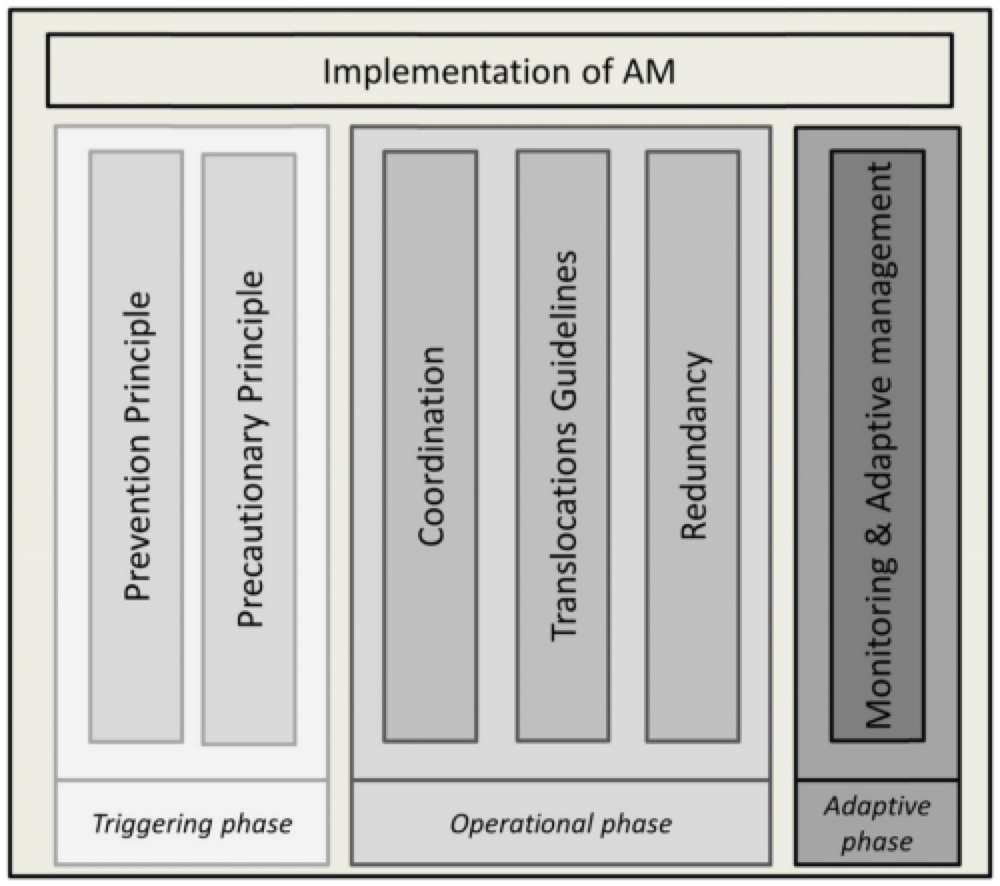
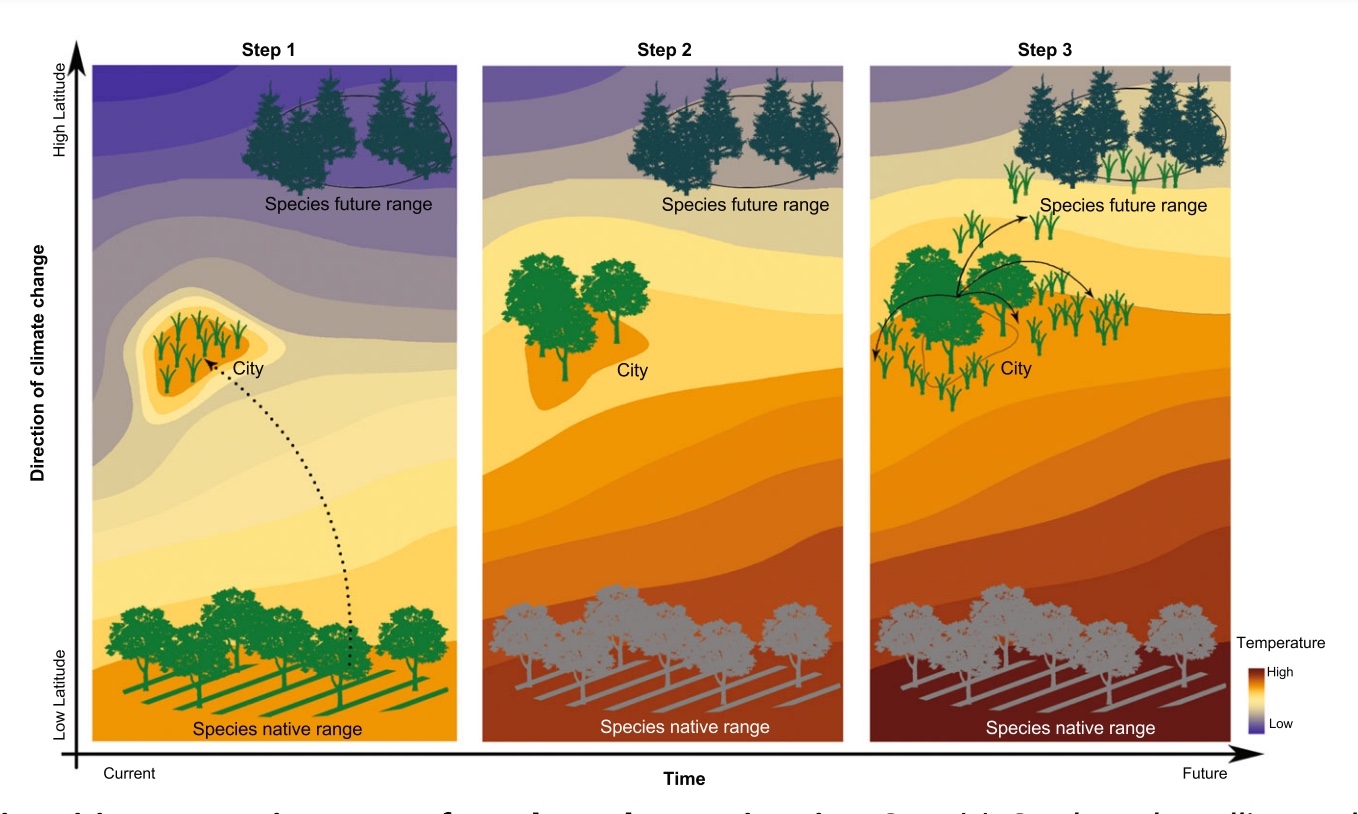
- by Guillame Peterson St-Laurent et al., 15 July 2019, Journal of Environmental Management
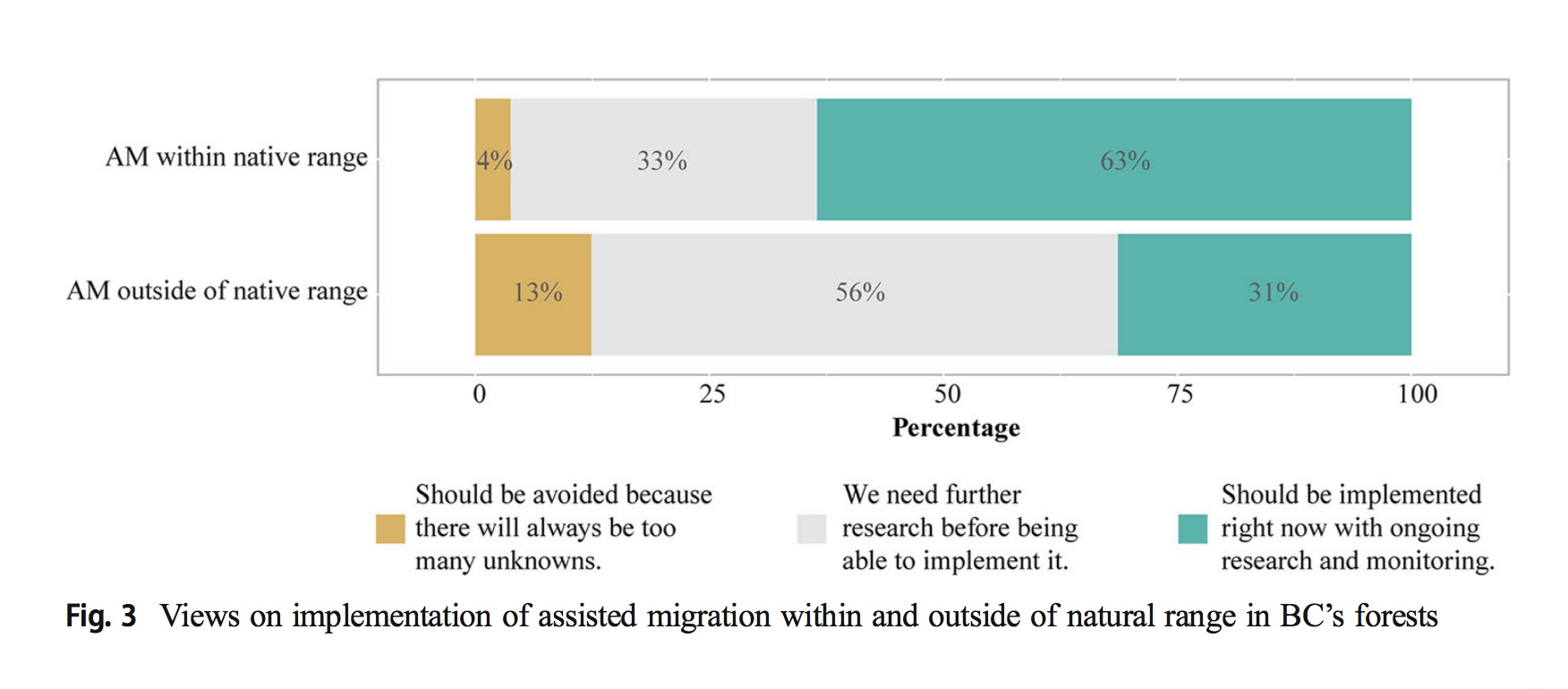
ABSTRACT EXCERPT: ... We use the example of genomics-based assisted migration (within and outside of natural range) in British Columbia's (BC) forests to examine the relative roles of and interactions between trust in different forestry actors and knowledge of forestry in shaping public support for this new and potentially controversial management alternative. Our results, based on an online survey, reveal low public trust in governments and the forest industry combined with low levels of public knowledge about forest management. We find that individuals who are more trusting of decision-makers and other important forestry actors hold higher levels of support for assisted migration. Higher levels of forestry knowledge are linked with support for assisted migration within native range, whereas no knowledge effect is observed for assisted migration outside of native range. We discuss the implications of these observations and provide recommendations to more fully engage with the challenges of low levels of trust and knowledge in this context.
Editor's note: In the Sci-Hub full text link, the term "assisted migration" appears 60 times in this article.
Should We Move the Whitebark Pine? Assisted Migration, Ethics and Global Environmental Change - by Clare Palmer and Brendon M.H. Larson, 2014, Environmental Values
Editor's note: This paper excels in exploring the range of values from the deep-ecology "intrinsic value" of a species and its "wildness" to human-centered aesthetic values and humans prioritizing the suffering of individuals of "sentient" species over the extinction of a non-sentient species.
ABSTRACT: Some species face extinction if they are unable to keep pace with climate change. Yet proposals to assist threatened species' poleward or uphill migration ('assisted migration') have caused significant controversy among conservationists, not least because assisted migration seems to threaten some values, even as it protects others. To date, however, analysis of ethical and value questions about assisted migration has largely remained abstract, removed from the ultimately pragmatic decision about whether or not to move a particular species. This paper uses the case study of the whitebark pine, a keystone species of sub-alpine habitats in western North America, to consider how particular cases of assisted migration may be ethically approached. After taking into account the value of species, wildness, place, ecosystems, culture and sentient animals, we conclude that, on balance, there appear to be good reasons to move the whitebark pine.
EXCERPTS: ... The assisted migration of rust-resistant strains of whitebark pine may be the only way to preserve the species for the future. If we think that a species has some kind of intrinsic value or moral status, this is a presumptive reason in favour of moving it. This reason might be defeated if the whitebark pine was likely to be invasive in its new location, and to threaten native species, but this is extremely unlikely to happen. Indeed, for around sixty years, until relocated whitebark pine starts setting significant numbers of cones, it will remain exactly where we planted it. Even then, it will only propagate with a healthy nutcracker [bird] population in situ. Invasiveness is thus not a plausible reason to ressist relocation. In addition, whitebark pines, even before they start to set seeds that may be a useful source for high-altitude animals and birds, could provide ecosystem services such as retaining soil moisture, modifying soil temperatures, slowing the progression of snowmelt and helping to moderate flooding at lower elevations...
The landscapes into which the whitebark pine would be relocated are not going to remain as they are. The species existing there will also be moving upward and poleward. New species will move in. Novel ecosystems will form (see Hobbs et al., 2013). Narratives of place will have to change, and human influence will extend across the changing landscape anyway. The relocation of the whitebark pine would not be the intrusion of an alien into a long-established and unchanging ecosystem, but a new member of a novel ecosystem with some potentially useful functions. ... Those who argue that species have moral status might deepen these narratives in terms of 'restitution' to species, such as the whitebark pine, that have been forced out of other locations by human activities. For while it's ethically difficult to make an argument that restitution or reparation can be directly owed to non-humans (with the possible exception of sentient animals), it may be possible to defend the view that the assisted migration of threatened species, where humans are responsible for the threat, is a kind of symbolic reparation.
To conclude: we are not arguing here for anything as strong as a duty to relocate the whitebark pine. We do, however, consider that there are good reasons to do so, and few very strong ethical objections. Our claim is that if rust-resistant strains of whitebark pine can be clearly identified, then depending upon the outcome of economic- and feasibility-studies, and consultation at particular sites, there are likely to be places where the relocation of whitebark pine is ethically desirable... We have sought to be comprehensive in our taxonomy and analysis of values, partly as a model for future studies that must similarly attend to pertinent socio-ecological complexity, to include both biological nuance (e.g., the association here between whitebark pine and the Clark's Nutcracker) and social importance (e.g., the cultural values of the whitebark pine). We hope that this analysis of the particular case of the whitebark pine will help to provide the basis for future (and better) studies and applications of assisted migration.
• September 2014 - "Time for Trees to Pack Their Trunks?", by Greg Breining, Ensia Magazine.
EXCERPTS: ... When it comes to assisted migration to adapt to climate change, caution has generally trumped action. "Most places in the world are doing research but they're very reluctant to move into practice," says Sally Aitken. "We sold people so well on the 'local is best' that it's a big change to all of a sudden say, 'Well you might want to get your seed from somewhere else.'"... "The crux of it is that species are at risk because of climate change," says Dov Sax, deputy director with the Institute for the Study of Environment and Society at Brown University and member of a scientific group that studied assisted migration
In some cases, the aim is to save a species with nowhere to go, stranded by elevation or manmade obstacles. In other cases, the concern is less for individual species than for the overall well-being of ecosystems. In northeastern Minnesota, for example, white spruce, paper birch and balsam fir may all but disappear in a warming climate. They will surely exist in profusion farther north. But can other tree species expand northward quickly enough to take their place? ... It may sound like a can't-lose. But in reality, assisted migration of trees is controversial — risky, some scientists say. For example, TNC offered to plant Michigan white pine on state and federal land in northeastern Minnesota, but both the U.S. Forest Service and state Department of Natural Resources declined, so the trees were planted on county land instead. "They are still, and rightly so, taking a conservative approach to moving things around," says Meredith Cornett.
... Some scientists and conservationists warn that mixing species will degrade "ecosystem integrity." But as Dov Sax points out, perhaps a fifth of the plant species in North America already hail from somewhere else. And the notion of a "balanced" ecosystem doesn't have much support in scientific evidence. "What we think of as ecosystems are transient — very few of the associations of species that we see now that make up an ecosystem would have been together at most points in time in the past," says Sax. "What we get is that maybe communities will be together for a few thousand years." Novel ecosystems — a hodgepodge of natives and exotics — can function, but how much worse or how much better than native assemblages is still a puzzle. "That's the kind of thing we don�t have a handle on," Sax says.
...When it comes to assisted migration to adapt to climate change, caution has generally trumped action. "Most places in the world are doing research but they're very reluctant to move into practice," says Aitken. "We sold people so well on the 'local is best' that it's a big change to all of a sudden say, 'Well you might want to get your seed from somewhere else.'"
In Europe "most conservationists are quite conservative (what's in a name?) when it comes to introducing or translocating species," Sebastiaan Van der Veken, director of the natural area Regionaal Landschap Kleine en Grote Nete in Belgium, wrote in an email. "Even reintroduction of recently disappeared species or short-distance translocation is highly discussed. Most scientists in Europe think (or hope) creating robust networks and corridors will make it possible for species to migrate themselves. They even fear that putting assisted migration on the agenda will restrain policy makers from investing in these ecological networks 'because we can simply move species around when necessary.'"
Meanwhile, as scientists debate, there's evidence that ambitious gardeners and commercial nurseries are stealing the march. Commercial plant nurseries are raising, selling and introducing hundreds of native forest plants far beyond their natural ranges — in effect pushing the northern limits of these trees, shrubs and wildflowers on average more than 600 miles, according to a study by Van der Veken. Not surprisingly, however, there is a bias toward pretty, popular and economically valuable species. "So it would be wrong to rely on these nurseries and gardeners alone to solve the biodiversity problem induced by climate change," Van der Veken wrote. "Scientists have an important role in making a framework for assisted migration, including screening of endangered species, translocation risk analysis and post-migration monitoring." Other scientists concur that the willy-nilly experimentation in the garden or backyard is unsuited to remaking something as long-lived and self-sustaining as a native forest.
"Under pressure: Conservation choices and the threat of species extinction" - by Robin Gretory et al., May 2021, Climatic Change
EXCERPTS: Shifts in species ranges and viability introduced by climate change are creating difficult challenges for scientists and citizens. In many cases, the seriousness of threats to endangered species is forcing policy makers and resource managers to consider novel species protection strategies, either to complement or replace existing conservation approaches. This paper seeks to deepen understanding of public views on a range of conventional and novel management initiatives designed to protect species under the threat of extinction, based on results from an online survey conducted in the USA and Canada.... Results generally support the conclusion that, so long as a clear rationale is provided, there exists surprisingly widespread support for the adoption of novel management approaches to save threatened or endangered species even if this requires more intensive genetic and transformational interventions that are costly or ethically challenging.... Such novel climate-adaptive options are not without controversy, due to the ethical issues and ecological risks to which they give rise. For instance, assisted migration involves concerns regarding the transgression of natural boundaries (Aubin et al. 2011; Hancock and Gallagher 2014; Peterson St-Laurent et al. 2018) and may contribute to the introduction of invasive species, pests, or diseases (Dumroese et al. 2015; Park and Talbot 2012; Pedlar et al. 2012). Objective setting by triage also raises moral issues in terms of deciding which species are conserved, how these decisions are made, and by whom (Bottrill et al. 2008; Knight et al. 2019). Because of the risks and values-based thresholds inherent in these more interventionist management responses, segments of the public as well as experts may prefer to rely on status quo or business-as-usual conservation strategies, at times favouring non-interventionist policies that step back and let nature take its course (Hagerman and Satterfield 2014).
"Responding to climate change in forest management: two decades of recommendations" - by Shannon M. Hagerman and Ricardo Pelai, October 2018, Frontiers in Ecology and Environment
EXCERPTS OF "ASSISTED MIGRATION" (the term or its cognates appears 4 times in the paper and twice in the references): ... Considering the pace of institutional change that characterizes many resource management contexts, it seems likely that amended rules and legislation to enable potential transformative, unconventional practices like assisted migration (Klenk 2015) will unfold slowly and may be controversial (Figure 1). This potential for dispute relates to the often-overlooked role of values and worldviews in shaping preferences for transformative policies and management actions (Moore et al. 2014; Andrachuk and Armitage 2015). Put simply, new policies may be rejected outright for values-based reasons (Baron and Spranca 1997; Hagerman and Satterfield 2013), such as the position that humans should (or should not) intervene in nature. We there- fore strongly agree with Andrachuk and Armitage (2015), who similarly asserted that values should be taken into account when evaluating potential pathways for transformation.... The analysis presented here provides a systematically derived synthesis of two decades of recommendations for forest management in response to climate change. We offer an empirical basis for future comparisons of recommendations through time, as well as a reference for comparisons between scientific and practitioner perspectives....
"Expert views on biodiversity conservation in an era of climate change" - by Shannon Hagerman et al., February 2010, Global Environmental Change
EXCERPTS (featuring "assisted migration"): Conservation scientists have responded with a range of adaptive conservation strategies. These include proposals for dynamic protected areas (Bengtsson et al., 2003; Rayfield et al., 2008) assisted migration (McLachlan et al., 2007; HoeghGuldberg et al., 2008; Richardson et al., 2009), and most prominently, the expansion of linked networks of static protected areas (e.g. Hannah et al., 2002; Hannah, 2008).... We pay particular attention to the interacting roles of (i) the history of debate about new policy proposals, (ii) the state of the science including uncertainty, (iii) the role of values and beliefs and (iv) policy windows. Below, we detail why these particular variables matter in this problem context and with respect to the purpose of this paper.
All interviewees expressed the view that a paradigm shift in conservation practice was required to adapt to the impacts of climate change.... Assisted migration the deliberate translocation of an imperiled species from one location to another where it has not existed in recent history, is a notable relative exception (e.g. McLachlan et al., 2007; Richardson et al., 2009).
... Overall, eight respondents initially expressed opposition to assisted migration interventions (six affiliated with NGO, one government and one academic). However, over the course of discussion, all but one respondent noted that interventions such as assisted migration would be necessary given the impacts of climate change. The tension surrounding discussions both about assisted migration and triage principles for management and objective setting arguably reflect different perspectives on 'nature' and the role of humans within/outside of 'it'. On the one hand, the statement above by Expert 17 reflects the view that true 'nature' is something 'out there', and separate from human influence. By this framing, all human manipulation is 'intervention', seen as unnatural and potentially unethical. On the other hand, the statements of Experts 2 and 13 above reflect the perspective that nature is a co-produced, integral and linked component of society. Viewed through the lens of linked SESs, so-called 'interventions' such as assisted migration can be viewed as part of the evolution of management and changing means and objectives through time (however undesirable this may seem and noting the need for great caution in potential implementation). Combined, these responses reveal how different perspectives on nature can shape expressed preferences for management, which have the potential to influence policy design and implementation.
April 2010 - "The Value of Species and the Ethical Foundations of Assisted Colonization",
by Ronald Sandler, in Conservation Biology.
"... Many species are imperiled by the accelerated rate ofclimate change, often in combination with other anthropogenic stressors, and frequently anthropogenic barriers
(e.g., urban and agricultural) prevent them from coping by shifting their geographical ranges (e.g., Hulme 2005; Parmesan 2006; Williams et al. 2007). Advocates of assisted colonization (or assisted migration) argue that helping species over these barriers is justified because it keeps them from going extinct, thereby preserving their value (e.g., Barlow & Martin 2004/2005; McLachlan et al. 2007; Hoegh-Guldberg et al. 2008). Therefore, the case for assisted colonization depends fundamentally on claims, both explicit and implicit, about the value of species. A complete discussion of assisted colonization needs to include assessment of these claims.... The paradigmatic cases of and candidates for assisted colonization discussed in the professional and popular literatures indicate. These cases include relocating the Quino checkerspot butterfly (Euphydryas editha quino) to higher elevations (e.g., Zimmer 2007; Hoegh-Guldberg et al. 2008; Marris 2008), the Florida torreya (Torreya taxifolia) to North Carolina (e.g., Barlow & Martin 2004/2005; Schwartz 2004/2005; Fox 2007), and the narrow-faced kangaroo rat (Dipodomys venustus) north of San Francisco (Berdik 2008). In each case, the aim is to prevent the species from going extinct, not to improve the ecological conditions of the recipient system. The ecological value of the species is not the basis for, and does not justify, the assisted colonization."
ABSTRACT: Discourse around assisted colonization focuses on the ecological risks, costs, and uncertainties associated with the practice, as well as on its technical feasibility and alternative approaches to it. Nevertheless,
the ethical underpinnings of the case for assisted colonization are claims about the value of species. A complete discussion of assisted colonization needs to include assessment of these claims. For each type of value that
species are thought to possess it is necessary to determine whether it is plausible that species possess the type of value and, if so, to what extent their possessing it justifies assisted colonization. I conducted such an assessment for each of the predominant types of value ascribed to species: ecological, instrumental (including option value), existence, and intrinsic value (including interest-based, objective, and valuer-dependent intrinsic
value). The vast majority of species, including several that have been proposed as candidates for assisted colonization, have much less value than is often presumed. Moreover, with respect to some types of value,
assisted colonization would not fully preserve the value of the target species even if it were to keep the target species in existence. Therefore, the case for assisted colonization is significantly weaker and more qualified than its advocates often suppose. There may be exceptional species for which assisted colonization is well justified�and for this reason, case-by-case assessment is necessary — but in general the burden of justification generated by the ecological risks associated with assisted colonization is not met by the value potentially preserved by assisted colonization. This suggests that assisted colonization ought to have, at most, a very minor role in the portfolio of ecosystem management practices, even as it pertains to species conservation under conditions of rapid climate change.
END OF VALUES SECTION (for assisted migration scholarly links)
"Agency launches long-term spruce, aspen treatment plan by Dennis Webb, The Daily Sentinel (Grand Junction, Colorado), 6 August 2016.
EXCERPTS: Responding to habitat shifts resulting from climate change will be one of the considerations for the Grand Mesa, Uncompahgre and Gunnison national forests as the Forest Service embarks on a new forest treatment project over the next eight to 12 years. Its new Spruce Beetle Epidemic and Aspen Decline Management Response project is a response to about 223,000 acres where spruce have died from beetle infestation on the forests, and 229,000 acres that have been affected by what's called Sudden Aspen Decline, over a decade.
The Forest Service expects mortality in spruce stands "to continue at relatively high levels for several years to come," according to the final environmental impact statement for the project. In 2009 the detection of new areas of aspen decline dropped considerably, but stands already affected continue to decline, and the Forest Service expects the aspen and spruce problems to be exacerbated in the future by climate change.
While the new forest treatment plan is intended to also address other goals like reducing safety hazards such as falling trees and increased wildfire danger, improving forest resiliency is a key goal. That includes trying to make the forest resilient in the face of a changing climate. "In the climate change world, that's called adaptation measures — basically trying to adapt the forest to a changing climate," said Jim Worrall, a Forest Service forest pathologist who helped do the climate modeling. He said quite a few outcomes of the management response project "could help adapt the forest to a warmer and potentially drier climate."
Worrall said where logging of beetle-killed spruce occurs, that could provide an opportunity for regenerating those acres with more aspen, or other trees more tolerant of an expected hotter, drier climate, such as Douglas fir, ponderosa pine and blue spruce. "I think at first it's going to be baby steps because people are a little bit cautious, and reasonably so, about completely changing a (forest) cover type," he said. Where there's spruce-fir forest, it might be crazy to start planting pinon-juniper now, even if models say it would be good pinon-juniper habitat by the end of the century, he said.
Also by Dennis Webb, 6 August 2016, "Cycle of decline: Estimate portends big changes in makeup of forests".
EXCERPTS: Lovers of the local high country could find a recent projection of a warming world's impacts on area forests to be chilling. By 2060, according to a U.S. Forest Service estimate, almost all of the Uncompahgre Plateau would no longer be able to sustain growth of new aspen and spruce, meaning that the plateau could be virtually aspen- and spruce-free by century's end after the remaining trees die. On the southern and eastern fringes of Grand Mesa, aspen also could see sizable losses of suitable habitat by 2060, with spruce habitat largely slipping into a threatened category across the mesa, meaning the future climate isn't favorable to sustaining it.
The modeling used by the Forest Service found that 52 percent of current aspen distribution across the forests would be in the lost habitat category by 2060, and 42 percent in the threatened category, "meaning it is conceivable that 94 (percent) of current aspen distribution may not continue into the next century," the Forest Service says in its final environmental impact statement for the project, released earlier this year. Aspen habitat generally would be lost at low elevations, especially on south-facing slopes, with the western West Elks also sharing in that habitat loss. Some of that habitat loss could be offset by newly emerging habitat at higher elevations. But Samantha Staley, a Forest Service planner, says while the climate may shift to support the species at a higher elevation, that doesn't mean that other ecological components necessary to support the species will be present. Some higher elevations may not be suitable thanks to things such as poor soil conditions or rocky scree slopes. The model projects a 22 percent loss of current spruce distribution, and that 58 percent of distribution will become threatened, meaning that 80 percent of current distribution may not continue into the next century.
The model is based on an assumption of a continuing warming trend on the forests. The statement says temperatures are expected to rise 5.4 to 7 degrees by 2040-60. Higher temperatures could foster more spruce beetle outbreaks, further stress trees because of increased drought and result in more damage from wildfire. "I think what those maps show is stunning,"Zukoski said. "I think those kind of maps are extremely helpful because they permit people to see in their areas, places that they care about in their backyards, what the world's going to be like for their kids and grandkids, and for themselves if they live long enough. I think giving people that picture over that longer term really helps them understand how dramatic the impacts of climate change could be if we don't work darn hard to get a handle on it."
Meanwhile, a 2006 model created by researchers led by Gerald Rehfeldt, who worked at the Forest Service's Moscow Forestry Sciences Laboratory in Idaho before retiring, projected a 47 percent drop in suitable spruce habitat in the western United States in the decade around 2060, and a 72 percent loss by 2090. The Forest Service statement says that for the forests, that model was rebuilt using local data, more "topographical predictors," newer global climate models and carbon scenarios, and higher-resolution climate data. Its resulting projections are an average from three climate models and three greenhouse gas emission scenarios. "These are models, which necessarily include some amount of error," Staley said. She said it's based on the best available information today, and the Forest Service understands the science will be a lot better in a decade. "We have to look at it as it's not the gospel, but it's the best available scientific information that we have today about where vegetation may be headed in the future," Staley said.
Jim Worrall, a forest pathologist with the Forest Service�s Rocky Mountain Region 2, which includes Colorado, said Rehfeldt worked with him and others to localize his methods, and they developed a model for 13 tree species in southwest Colorado, including spruce and aspen. Worrall said one of the biggest uncertainties pertains to the climate models, because there are so many models and carbon scenarios to choose from. He said the forest-habitat predictions the Forest Service came up with concern him and he hopes they're wrong, but they're the best idea researchers now have of what the future holds. "It's really a very objective process. There's really no subjectivity in the model development," he said. Interpreting the results to make them simple and easy to digest can be a little subjective, he said. "But we've been pretty conservative in that," he said, adding that the predictions are based on what the models are telling researchers, and those models are built on a lot of data.
Staley said the agency is compelled to use that science in its planning and efforts to manage sustainably into the future, and current research acknowledges that warming will result in shifting of not just animals but trees in terms of habitat. "The environment is changing and that's why we're using as much information as we can to make the best decisions," she said. Forests are always changing, thanks to factors such as insect infestation and wildfire, and cycles such as aspen thriving first in disturbed areas and later being succeeded by other types of trees. But now the Forest Service is learning more about how that's happening in the context of "pretty rapid climate change," and what the forest may look like in that context, Staley said.
While every generation sees a slightly different version of a forest, future forests may be ones that people have never seen locally, she said. Rather than a mid-elevation forest shifting back and forth from aspen to spruce, it may shift to oakbrush. "And that's a new shift," she said. Oakbrush and mixed-mountain shrubs cover about a quarter of the Uncompahgre Plateau now.
[Jim Worrall] said of the modeling's findings, "I think we need to plan for the worst and hope for the best, is what I suggest. But most likely even if the models are a little off, we're going to be looking at very different conditions in the future for our children and our children's children than we have now."
"The assisted migration of western larch in British Columbia: A signal of institutional change in forestry in Canada? by Nicole L. Klenk and Brendon M.H. Larson, Global Environmental Change 2014.
EXCERPTS: Based on 46 interviews with policy actors across Canada, our results suggest that the deployment of the first assisted migration policy in Canada successfully avoided the controversy surrounding the idea in the scientific community by changing the scientific discourse associated with best forest management practices. The shift from an ecological discourse to a genetics discourse over forest policy in British Columbia signals what we might expect in future forest adaptation policy development in Canada.
Clearly, a genetics characterization of forests looks back to the distant past for guidance in future 'climate change fitness of species'. This represents a major conceptual shift, which significantly affects the temporal and spatial scales on which forest management planning occurs. This shift can be characterized, furthermore, by the deployment of new normative goals for forest management: rather than trying to recreate current forest composition and functions, the values and norms guiding forest management from a genetics perspective seek to accelerate forest transition to the future, to a 'climate resilient' state. This normative shift has real material consequences, such as enabling the movement of western larch 1000 km north of its current distribution, sidestepping the issue of its ecological appropriateness in the recipient ecological community, because from a 'climate fitness' perspective, western larch may eventually migrate to northern BC.
Thus given the genetical frame structuring the AM policy coordinative discourse, we argue that there is a major shift in the cognitive and normative content of ideas at the program level. However, when we analyzed the communicative discourse on western larch AM policy, we found that concerted efforts were taken by policy developers to downplay the significance of this program change in their communication to the public. To begin, educational and training outreach activities stressed the low risk associated with population range expansion, as the following interviewee explains: "We are very open and inclusive in talking about AM. I talk about the risk of action and inaction and more importantly the difference between the various forms of AM. We are very clear that we are not testing or interested in exotic translocations."
To sum up, our analysis suggests that policy analysts, forest geneticists and policy implementers in the BC Tree Improvement Branch characterized forests so as to highlight and diffuse an evolutionary theory of forest ecosystems that is tied to a new set of norms and values associated with a humanistic philosophy that are highly controversial in the conservation community—yet which, in turn, are signals of major institutional change in forest policy in BC. . . Our results suggest that the deployment of the first AM policy in Canada has successfully avoided the philosophical debates on AM in the conservation scientific community by changing the scientific discourse associated with best forest management practices (i.e., from an ecological point of view to a genetics point of view) and this discursive shift may signal what we might expect in future forest adaptation policy development in Canada.
Editor's note: See also Nicole L. Klenk, 2015, "The Development of Assisted Migration Policy in Canada: An Analysis of the Politics of Composing Future Forests", published in Land Use Policy. For a superb example of Canadian foresters utilizing the best communication skills in reported news stories, see the 14 June 2015 "Canadian scientists help trees adapt to changing climate". Here is the lead quote by researcher Sally Aitken, "Trees are adapted to historical climate and the climate's moving out from under them. We're using genomics to generate answers more quickly than they can." Here is her closing quote: "We have changed things to the point where we really have to foster the future of the environment and the forest. I think it would be very foolish and irresponsible to say, 'Let nature take care of itself.'" As well, see Aitken quoted in 29 January 2014 news story: "New genes for old forests as Canada warms": "So my research is focused on the best way to better match trees with new and future climates, to assist the movement of that genetic material through reforestation." Aitken is also quoted here (for Whitebark Pine, 18 September 2014, NYT): "For Trees Under Threat, Flight May Be Best Response".
See also; "Opinions on strategies for forest adaptation to future climate conditions in western Canada: Surveys of the general public and leaders of forest-dependent communities", 2014, Reem Hajjar et al., Canadian Journal of Forest Research.
REVIEW ARTICLE IN FORESTRY ADAPTIVE MANAGEMENT 2019: "Adaptation strategies and approaches for forested watersheds", 2019, by P. Danielle Shannon et al. [all 8 authors are associated with Northern Institute of Applied Climate Science] in Michigan, Climate Services, February 2019.
Editor's note: This 2019 review paper lays out the adaptive management process developed by NIACS over the years and reports examples of two user-groups in Massachusetts and Minnesota, only one of which (Knife River adaptation demonstration in Minnesota) chose to use elements of "assisted migration" as a management tool. Ricciardi and Simberloff 2009 is cited multiple times for its cautionary perspective, "Assisted Colonization Is Not a Viable Conservation Tool." It is unusual for a forestry paper to give this degree of credence to a perspective grounded not in traditional forestry but in conservation biology. On the plus side of using research to support actual and pragmatic user decisions, the NIACS approach in advising landowners on forestry climate adaptation is grounded in assisting management discussions on climate risk, management values and goals, and site-specific tactics — without staff or advisors making any recommendations along the way. This summary of what the group has been doing over the years is highly recommended. Pages 6-7 expressly deal with assisted migration
as one possible tactic for climate adaptation.
EXCERPTS: ... The overarching goal of this work is to help individuals and organizations within a diverse management community clearly articulate their decisions and how they have intentionally considered climate change, risk, and adaptation in their management.... Two major challenges in the practice of climate adaptation are (1) translating broad adaptation concepts (Millar et al., 2007) to specific, tangible actions (Swanston et al., 2016), and (2) clearly and explicitly identifying the adaptation intent of an on-the-ground action. We de- veloped an adaptive management process and step-by-step Adaptation Workbook in 2012 (Swanston and Janowiak, 2012; Swanston et al., 2016) that helps practitioners identify adaptation actions that address perceived climate threats and opportunities in pursuance of their stated management objectives. We also developed a series of adaptation menus that help people link their actions to broader adaptation stra- tegies that align with their values and objectives, generally define success, and explicitly identify intent (Swanston et al., 2016). Each menu addresses a different resource area, using relevant and appro- priate terms, strategies, approaches, and example tactics. Although the menu concepts focus on adaptation to climate change, the ideas are rooted in fundamental principles of natural resources management (Creed et al., 2011) and may also provide co-benefits to various conservation efforts and greenhouse gas mitigation goals. Critically, the menus are not guidelines and do not make recommendations. They instead represent numerous approaches to resource management that are often complementary, but always chosen by the user as they deem appropriate.... Importantly, the Adaptation Workbook and menus do not compel or influence natural resource managers to change their ecosystems or adopt new practices; instead they help mangers make intentional, climate-informed decisions best suited to their objectives, constraints, and perception of climate risks and opportunities.
... Strategy 4: Facilitate forest ecosystem adjustments through species transitions Approach 4.1: Favor or restore native species that are expected to be adapted to future conditions. Approach 4.2: Establish or encourage new mixes of native species. Approach 4.3: Disfavor species that are distinctly maladapted. Approach 4.4: Introduce species that are expected to be adapted to future conditions. Approach 4.5: Move at-risk species to locations that are expected to provide habitat.
"What is Novel About Novel Ecosystems: Managing Change in an Ever-Changing World", 2015, by Amy Truitt et al, Environmental Management
Editor's note: This 2015 paper is quoted at length because it places "assisted migration" within a tri-fold management regime which the future will necessarily bring to ecological restoration and conservation biology.
EXCERPTS: While over the past several hundred years most landscapes have been altered by anthropogenic activities, and in many cases outright habitat destruction, the terminologydescribing the resulting ecosystems is inconsistent and inadequate for effective cross-sectoral management. Due to the dynamic nature of ecosystems and the varied disciplines that have evolved in the Anthropocene to study them, it has proven challenging to find a universal language that defines 'novel' ecosystems (e.g., Milton 2003; Hobbs et al.2013;Morse et al.2014; see Table1). Equally challenging is the development of a common set of metrics to quantify ecosystem changes and the functional impacts resulting from those changes (Murcia et al.2014). Neutral, unambiguous, and consistent terminology with a framework for categorizing novel ecosystems is needed to classify, describe, and manage these systems. Such classification can facilitate decision-making in a landscape of diverse management objectives and variable social perceptions around novel systems (but see Murcia et al. 2014).
Priorities for and approaches to managing novel ecosystems vary based on perspective of the scientists, managers, and policy-makers along the anthropocentric�biocentric continuum. Practitioners with a biocentric perspective are more concerned with impacts that novel ecosystems have on biodiversity, community composition, and ecosystem function, while those with an anthropocentric perspective may be more interested in how novel ecosystems will affect ecosystem services on which humans depend. Management approaches to novel ecosystems range from strategies that actively promote the ability of novel ecosystems to achieve conservation goals (Zedler et al.2012), to active prevention of novel ecosystems (Stromberg et al.2009). These approaches range from accepting irreversible landscape alterations and promoting and accounting for ecosystem services provided by novel assemblages (Melo et al.2013) to active prevention of native species and ecosystem loss (Willis et al.2010; Lindenmayer et al.2008) or land restoration to prioritize ecosystem functions and processes in certain novel ecosystem scenarios (Hobbs et al.2009). On some sites, assisted migration, such as purposeful ymoving species to more climate suitable habitat (Richardson et al.2009) has been recommended. Some practitioners suggest that novel ecosystems should be recognized as achieving conservation goals and remain where they arise, rather than be managed (Hobbs et al.2006;2011; Martinet al.2012). Others suggest eliminating the term 'novel ecosystems' as it could be a hindrance to ecologically sound restoration efforts (see Murcia et al.2014). Acknowledging the diverse perspectives may facilitate compromise and development of mutually acceptable management approaches. The term itself may be useful in delineating to the public and policy-makers the far-reach-ing effects of anthropogenic activities on proximal and remote ecosystems (e.g., Blight and Ainley 2008; Holt-grieve et al.2011), which may facilitate greater consensus on appropriate management strategies.
Here we ask whether novel ecosystems are, in fact novel, or whether these systems are a manifestation of ongoing evolutionary change in ecosystem and species assemblages. Given the many definitions of novel ecosystems, we present previous and current definitions, offer a working definition for use in management decision-making, and differentiate the frameworks surrounding each to organize possible management approaches to novel ecosystems. Finally, we propose a categorization scheme and framework (Figs.1,2) for managing novel ecosystems now and in the future.
We introduce a framework for identifying how to manage novel ecosystems with three alternative directives to address the spectrum of challenges posed by these ecosystems. The directives include managing against, tolerating, and managing for novel ecosystems and can be directly tied to management objectives (Fig.2). We provide examples and discuss the social, economic, and ecological advantages and disadvantages of each.
There is a threshold in some affected ecosystems at which point either managing against or tolerating is no longer an option (Fig.1). Although this alternative is less common and may be more controversial, we expect the need to manage for novel ecosystems and the services they have evolved to provide will become more prevalent in the future under changing climatic conditions and increased globalization. Where species have been lost or have severely declined and restoration has been unsuccessful, or where land use change has been intensive, managers may find that managing for novel ecosystems provides the only or best alternative. . . Other examples of managing for novel ecosystems include the plethora of introduced plant species that have become important in the agricultural sector; assisted migration of more climate-suitable tree species to managed forests (Gray et al.2010; Duveneck and Scheller 2014); managing non-native species assemblages in parks (e.g., Underwood et al.2004); non-native species used for recreational purposes (e.g., Crawford and Muir 2008; Britton et al.2011) and non-native species for conservation (Schlaepfer et al.2010; but see Vitule et al.2012for rebuttal).
"The Fight to Unmuzzle Canada's Scientists" by Stephen Buranyi, Motherboard 27 August 2015
EXTRACTS: Dr. Phil Burton's work was also far outside what he thought the government might consider hot button. Dr. Burton, a professor at the University of Northern British Columbia, had previously spent 20 years with the Canadian Forest Service studying boreal forests in northern Canada. He recalls getting permission from Ottawa to attend a conference, but his permission came with a paradoxical request: "The conference was a series on disturbance dynamics in boreal forests, and the theme that year was climate change. I was told not to talk about climate change or forestry impacts. Well that's the whole point of the conference!" he said. . . "I work with government scientists every day, literally every single day, and they've just had their legs completely cut out from under them," said Alana Westwood, a graduate student in biology at Dalhousie University.
The Liberal Party of Canada has indicated it's interested in staking its claim early as the party of science. In May, Liberal MPs introduced a motion to "un-muzzle" Canadian scientists and introduce a parliamentary science officer, and a month later Liberal Leader Justin Trudeau announced that if elected he would review some of the legislative changes to environmental acts under Harper. The Liberal motion was largely symbolic; the Conservative majority sunk it fast. But it was a start, and the campaign promises remain on record.
Editor's note: A few months after this report was published, the Liberal Party did become the majority party, and Justin Trudeau did oust Stephen Harper from the office of Prime Minister. The muzzles are gone!
"Conservation Biology: The End of the Wild" by Emma Marris, Nature 12 January 2011, 469:150-52
"Climate change means that national parks of the future won't look like the parks of the past. So what should they look like? . . . [National Park Service director Jonathan] Jarvis has suggested the possibility of moving species outside their native ranges to give them a better chance of surviving — just not right away. "The big point here is that we are willing to face these questions," he says. "We are not afraid to talk about them."
SEE ALSO an Dec 2010 online interview with NPS Director Jonathan Jarvis on climate change and park management, including "assisted migration".
"Defining Migration" chapter of Brian Keel's PhD thesis.
Brian Keel intentionally coined the term "assisted migration." This short chapter will be useful for those engaged in considering whether "assisted migration" or "assisted colonization" is the best term for the kinds of conservation actions now beginning to be considered. Editor's note: The historical sequence of controversy over terminology can be accessed here: "Assisted Migration or Assisted Colonization: What's in a Name?"
"Assisted Colonization: CBC Radio Interview"
A terrific AUDIO exploration of the controversy, which aired 24 July 2008. Part 1 is the supportive side, via an interview with Prof. Camille Parmesan. Part 2 is an interview with an invasive species researcher that is very critical of the idea. Part 3 is a not-to-be-missed radio spoof of the idea.
"Tending to Our Rambunctious Garden" Q&A with journalist Emma Marris, OnEarth 28 September 2011
Emma Marris's 2011 book, Rambunctious Garden, has "Assisted Migration" as the topic and title for chapter 5. The Q&A linked above is an excellent short introduction to the reach of this book in highlighting the shifting norms and values in conservation biology and land management today. Assisted migration advocate Connie Barlow posted a positive review of the book on Amazon: "Rewilding, Assisted Migration, Ecological Restoration, and More". There's also an excellent dot-Earth video interview of Marris on youtube.
"The Age of Man Is Not a Disaster" - Op-ed by Emma Marris, Peter Kareiva, Erle C Ellis, New York Times 7 December 2011
EXCERPT: "We can accept the reality of humanity's reshaping of the environment without giving up in despair. We can, and we should, consider actively moving species at risk of extinction from climate change."
"Guardian Angels" article by Janet Marinelli, Audubon Magazine, May/June 2010.
In-depth exploration of "the biggest controversy in contemporary conservation science." Engagingly written for both a popular and professional audience, journalist Marinelli draws from her interviews with leading scientists, horticulturalists, and activists to present the core arguments for and against assisted migration. A site visit to an endangered plant breeding facility (the Atlanta Botanical Garden) is paired in the article with Marinelli's eye-witness description of "eco-vigilante" action, when the loose-knit citizens group Torreya Guardians intentionally planted into forested landscapes of mountainous North Carolina 31 seedlings of the highly endangered Florida Torreya — an assisted migration of some 400 miles northward of historically known native habitat.
"Taking Wildness in Hand: Rescuing Species" article by Michelle Nijhuis, Orion Magazine, May/June 2008.
A lengthy and elegant feature article that explores the human side of the controversy over assisted migration, with Torreya taxifolia providing the focal point, pro and con, and with actions by the citizen group Torreya Guardians